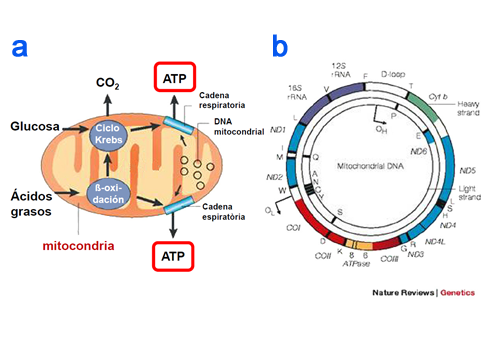
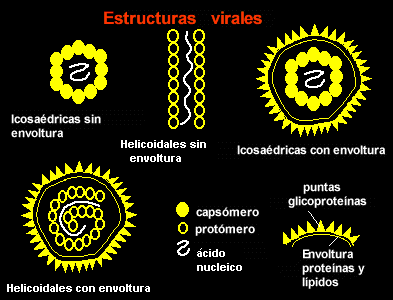
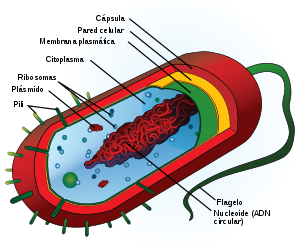
Se denomina eucariotas a todas las células que tienen su material hereditario fundamentalmente (su información genética) encerrado dentro de una doble membrana, la envoltura nuclear, que delimita un núcleo celular.Las células eucariotas son las que tienen núcleo a lo contrario que las procariotas que carecen de el. Se llaman las célulastontas a aquellas que solo tienen medio núcleo y la mitad de su materia genética esta dispersada por toda ella. A los organismos formados por células eucariotas se les denomina eucariontes.
En los organismos eucarióticos,
la mayoría de los genes se encuentra en los cromosomas del núcleo.
Las especies
eucarióticas se clasifican como:
Diploides:
(Dos series de cromosomas en el núcleo).
Haploides: (Una sola serie de cromosomas en el nucleo), la mayoría de
algas y hongos son haploides y el resto de eucariotas (incluyendo plantas y
animales) son diploides.
La letra n se utiliza para
designar el número de cromosomas de un genoma nuclear de un organismo.
Diploide: 2n.
Haploide: n.
Las condiciones 3n, 4n, 5n,: Se conocen como poliploides.
En una célula diploide, puesto
que hay dos series cromosomitas, existen dos cromosomas de cada tipo. Los
miembros de cada par se conocen como cromosomas
homologos.
Los miembros de un par homologo básicamente son identicos y
aportan los mismos genes, aunque muchas veces diferentes alelos (formas diferentes de un mismo gen).
Para determinar el número de cromosomas
de una célula, estos simplemente se tiñen y se cuentan al microscopio óptico.
3.2.1 ADN LINEAL Y EMPAQUETAMIENTO
Una célula humana contiene
alrededor de 2 metros de DNA (1
metro por cada serie cromosomica). El cuerpo humano está
constituido por unas 1013 células y cada célula es diploide, por lo
que contiene en total unos 2x1013 metros de DNA.
La distancia de la tierra al sol
es 1,5 x 1011 metros…
Ello significa que el DNA de
nuestro cuerpo podría extenderse hasta el sol y volver casi 100 veces. Este
hecho significa que el DNA de las eucariotas debe estar empaquetado de una
forma muy eficaz.
3.2.1.1 HISTONAS
De hecho el empaquetamiento
ocurre en el núcleo donde el DNA se condensa en 46 cromosomas, todo ello en un
núcleo de 0,006 mm
de diámetro! Para entender como ocurre esto hay que conocer la estructura
tridimensional de los cromosomas.
Al microscopio los cromosomas
aparece como fibras de 30 nm de diámetro, o que indica que la molécula de DNA
debe estar muy plegada.
Histonas: La mezcla completa de materiales de los que se compone el cromosoma se conoce
como cromatina. Se trata de DNA y
proteínas.
Las histonas son proteínas
asociadas al DNA en los nucleososmas. Los nucleosomas (10 nm) están formados
por un octamero compuesto por dos unidades de cada una de las histonas (H2A,
H2B, H3 Y H4).
3.2.1.2 SOLENOIDES
El DNA (asociado a las histonas)
da dos vueltas alrededor de cada octamero de los nucleosomas, el
nucleosoma es una forma distendida, de una forma muy enrollada denominado solenoide (30 nm) , el solenoide mantiene su forma mediante otra histona H1.
Para conseguir el primer nivel de
empaquetamiento el DNA se enrolla alrededor de las histonas, que actúan, como
bobinas de un hilo, un nuevo enrollamiento genera la conformación del
solenoide.

3.2.3 CROMOSOMAS
Los cromosomas se encuentran muy enrollados, si
un solenoide tiene de diámetro 30 nm, un cromosoma condensado tiene 700 nm.
Para esto el solenoide se enrolla sobre un esqueleto proteico compuesto de la
enzima topoisomerasa II, que es capaz de pasar una cadena de DNA a través de otra
.
En los niveles sucesivos de
empaquetamiento cromosómico:
1) El DNA se enrolla sobre los nucleosomas que
actúan como bobinas de hilo.
2) Los nucleosomas se enrollan formando un
solenoide.
3) El solenoide forma lazos anclados a un esqueleto central.
4) El
esqueleto unido a los lazos se dispone como un solenoide gigante.
Bibliografía:
Wu RS, Panusz HT, Hatch CL y Bonner WM (1986) Histones and their modifications, Crit Rev Biochem 20: 201-263.
http://www.ucm.es/info/genetica/grupod/Cromoeuc/cromoeuc.htm